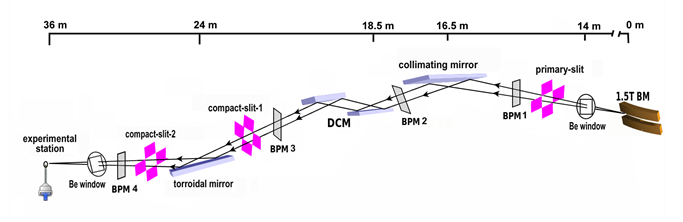
The protein crystallography (PX-BL21) beamline has been designed for macromolecular crystallography i.e. diffraction studies on single crystals of protein, DNA and their complexes. It has also been designed to perform high-pressure studies using single crystal and powder diffraction experiments. The beamline can be tuned to desired energy in the range between 5 to 20 keV (with bandwidth of ~1 eV) corresponding to wavelength of 2.48– 0.62 Å. The beamline is equipped to perform Single- and Multi- wavelength anomalous diffraction experiments (SAD, MAD) by fine-tuning energy of beam with required energy resolution. The typical time for data acquisition is about 5 to 30 sec per frame depending on the crystal quality and synchrotron beam current. The beamline is associated with a well-equipped biochemical laboratory and cold rooms to grow, cryo-soak and cryo-freeze the protein crystals
A panoramic view of BL21
A beamline view in Hutch-1
Beamline parameters & Optical layout
Beamline name
PX-BL21
Source type
1.5 T bending magnet
Mirrors
Both CM and TM are 1.2 m single-crystal Si with Rh coating
Monochromator
Water-cooled DCM with Si (111) or Si (220) crystals
Energy range (keV)
5–20
Wavelength range (A°)
0.62–2.48
Beam size (μm) (FWHM)
500 x 500
Flux (photons s-1) *
1.2 x 1010
Schematic of PX-BL21 beamline
Experimental station
Experiments
MONOCHROMATIC, SAD, MAD
Experimental stage
MARdtb
Goniometer
Single-axis MARdtb
Detector type
CCD
Detector
MARCCD 225 (Rayonix), 225mm x 225mm (active area)
Detector resolution
3072 x 3072 pixels; pixel size: 73 x 73 μm2
Detector saturation counts
60,000 photons/pixel
Detector angular displacement (2ϴ)
0–30°
Fluorescence detector
AMPTEK XR 100 SDD
Cryo sample cooler
Oxford Cryostream - 700 series
Cryogenic Robotic sample changer
Carousel of 19 samples (MAR research)
Biochem Lab Facility
Well-equipped working laboratory with cold rooms
Cryo-pins recommended
SPINE
On-site data processing server
HP Z800 (12-core Xeon 2.67 GHz, 24 GB RAM)
Software
Automar, XDS, Mosflm, CCP4, PHENIX, Coot, Pymol
Experimental facility
Macromolecular diffraction at fixed Single wavelength (0.62–2.48 Å)
Single-wavelength anomalous dispersion (SAD) on macromolecular crystals
Multi-wavelength anomalous dispersion (MAD) on macromolecular crystals
Single crystal diffraction of larger unit cell crystals (>10 Å) of organic molecules
Biochemical & crystallization facility
The PX beamline is associated with a well-equipped biochemical laboratory and cold rooms. This laboratory has all the necessary infrastructure and facility for protein expression, purification and crystallization. It also has dedicated crystal growth rooms, incubators and microscopes. The users may carry out necessary post-crystallization soaks and crystal freezing using this facility. There is a LN2 Dewar to store and maintain the crystals under the liquid nitrogen temperature.
How to apply for beam-time at PX-BL21?
The beam-time is allotted to users through an online portal (https://www.info-rrcat.ernet.in/beamline/). However, it is requested to users (prior to online application) to send an email to the contacts (ravimakde@rrcat.gov.in) for knowing the availability of the beamline/synchrotron with the proposed dates of their experiments. Users are requested to arrange their travel only after they receive the confirmation from the beamline team.
How to acknowledge the use of the beamline?
The use of PX-BL21 for data collection and/or testing of the crystals is requested to be acknowledged by both, a) citing the beamline paper, and b) using the following phrase in the acknowledgement "PX-BL21 beamline (BARC) at Indus-2, RRCAT, Indore".Beamline paper: Ashwani Kumar, Biplab Ghosh, H. K. Poswal, K. K. Pandey, Jagannath, M. V. Hosur, Abhilash Dwivedi, Ravindra D. Makde and Surinder M. Sharma, "Protein crystallography beamline (PX-BL21) at Indus-2 synchrotron, J. Synchrotron Rad. (2016). 23, 629-634. doi: http://dx.doi.org/10.1107/S160057751600076X
Application Areas
Solving high-resolution crystal structure of protein, DNA, RNA or their complexes
Solving high-resolution crystal structures of protein bound to different ligands
Solving crystal structure of organic molecules with larger unit cell
1.
Kumar A, Singh R, Ghosh B, Makde RD.
Crystal structure of aspartyl dipeptidase from Xenopus laevis revealed ligand binding induced loop ordering and catalytic triad assembly.
Proteins. 2022 Jan;90(1):299-308
1.
Parihar PS, Singh A, Karade SS, Sahasrabuddhe AA, Pratap JV.
Structural insights into kinetoplastid coronin oligomerization domain and F-actin interaction.
Curr Res Struct Biol. 2021 Oct 14;3:268-276.
2.
Afsar M, Shukla A, Kumar N, Ramachandran R.
Salt bridges at the subdomain interfaces of the adenylation domain and active-site residues of Mycobacterium tuberculosis NAD+-dependent DNA ligase A (MtbLigA) are important for the initial steps of nick-sealing activity.
Acta Crystallogr D Struct Biol. 2021 Jun 1;77(Pt 6):776-789.
3.
Kumari P, Sundaram R, Manohar K, Vasudevan D, Acharya N.
Interdomain connecting loop and J loop structures determine cross-species compatibility of PCNA.
J Biol Chem. 2021 Jul;297(1):100911. doi: 10.1016/j.jbc.2021.100911. Epub 2021 Jun 25. PMID: 34175309; PMCID: PMC8319368.
4.
Pandey B, Dev A, Chakravorty D, Bhandare VV, Polley S, Roy S, Basu G. Insights on the disruption of the complex between human positive coactivator 4 and p53 by small molecules. Biochem Biophys Res Commun. 2021 Nov 12;578:15-20. doi: 10.1016/j.bbrc.2021.09.020. Epub 2021 Sep 10. PMID: 34534740.
5.
Vajravijayan S, Pletnev S, Luo Z, Pletnev VZ, Nandhagopal N, Gunasekaran K.
Crystallographic and calorimetric analysis on Pleurotus ostreatus lectin and its sugar complexes - promiscuous binding driven by geometry.
Int J Biol Macromol. 2020 Jun 1;152:862-872.
6.
Gupta GD, Bansal R, Mistry H, Pandey B, Mukherjee PK.
Structure-function analysis reveals Trichoderma virens Tsp1 to be a novel fungal effector protein modulating plant defence.
Int J Biol Macromol. 2021 Nov 30;191:267-276. doi: 10.1016/j.ijbiomac.2021.09.085. Epub 2021 Sep 20. PMID: 34547313.
7.
Shukla A, Afsar M, Kumar N, Kumar S, Ramachandran R.
Structure based identification of first-in-class fragment inhibitors that target the NMN pocket of M. tuberculosis NAD+-dependent DNA ligase A.
J Struct Biol. 2021 Mar;213(1):107655.
8.
Chakraborty S, Biswas M, Dey S, Agarwal S, Chakrabortty T, Ghosh B, Dasgupta J. The heptameric structure of the flagellar regulatory protein FlrC is indispensable for ATPase activity and disassembled by cyclic-di-GMP. J Biol Chem. 2020 Dec 11;295(50):16960-16974.
9.
Chakrabortty T, Roy Chowdhury S, Ghosh B, Sen U.
Crystal Structure of VpsR Revealed Novel Dimeric Architecture and c-di-GMP Binding Site: Mechanistic Implications in Oligomerization, ATPase Activity and DNA Binding.
J Mol Biol. 2021 Nov 10;434(2):167354.
10.
Viswanathan V, Rani C, Ahmad N, Singh PK, Sharma P, Kaur P, Sharma S, Singh TP.
Structure of Yak Lactoperoxidase at 1.55 Å Resolution.
Protein J. 2021 Feb;40(1):8-18.
11.
Singh PK, Pandey S, Rani C, Ahmad N, Viswanathan V, Sharma P, Kaur P, Sharma S, Singh TP.
Potassium-induced partial inhibition of lactoperoxidase: structure of the complex of lactoperoxidase with potassium ion at 2.20 Å resolution.
J Biol Inorg Chem. 2021 Feb;26(1):149-159.
12.
Sundaram R, Manohar K, Patel SK, Acharya N, Vasudevan D.
Structural analyses of PCNA from the fungal pathogen Candida albicans identify three regions with species-specific conformations.
FEBS Lett. 2021 May;595(9):1328-1349.
1.
Karade SS, Ansari A, Srivastava VK, Nayak AR, Pratap JV.
Molecular and structural analysis of a mechanical transition of helices in the L. donovani coronin coiled-coil domain.
Int J Biol Macromol. 2020 Jan 15;143:785-796.
2.
Agrawal R, Goyal VD, Singh R, Kumar A, Jamdar SN, Kumar A, Makde RD.
Structural basis for the unusual substrate specificity of unique two-domain M1 metallopeptidase.
Int J Biol Macromol. 2020 Mar 15;147:304-313.
3.
Singh AK, Datta A, Jobichen C, Luan S, Vasudevan D.
AtFKBP53: a chimeric histone chaperone with functional nucleoplasmin and PPIase domains.
Nucleic Acids Res. 2020 Feb 20;48(3):1531-1550.
4.
Singh R, Deshmukh S, Kumar A, Goyal VD, Makde RD.
Crystal structure of XCC3289 from Xanthomonas campestris: homology with the N-terminal substrate-binding domain of Lon peptidase.
Acta Crystallogr F Struct Biol Commun. 2020 Oct 1;76(Pt 10):488-494.
1.
Sharma M, Kumar V.
Crystal structure of BinAB toxin receptor (Cqm1) protein and molecular dynamics simulations reveal the role of unique Ca(II) ion.
Int J Biol Macromol. 2019 Aug 23. pii: S0141-8130(19)35380-2
2.
Megta AK, Mishra AK, Palva A, von Ossowski I, Krishnan V.
Crystal structure of basal pilin SpaE reveals the molecular basis of its incorporation in the lactobacillar SpaFED pilus.
J Struct Biol. 2019 Jul 1;207(1):74-84. doi: 10.1016/j.jsb.2019.04.016.
3.
Singh AK, Datta A, Jobichen C, Luan S, Vasudevan D.
AtFKBP53: a chimeric histone chaperone with functional nucleoplasmin and PPIase domains.
Nucleic Acids Res. 2019 Dec 6. pii: gkz1153. doi: 10.1093/nar/gkz1153.
4.
Karade SS, Ansari A, Srivastava VK, Nayak AR, Pratap JV.
Molecular and structural analysis of a mechanical transition of helices in the L. donovani coronin coiled-coil domain.
Int J Biol Macromol. 2019 Nov 25. pii: S0141-8130(19)31056-6. doi: 10.1016/j.ijbiomac.2019.09.138.
5.
Pal K, Yadav M, Jain S, Ghosh B, Sen R, Sen U.
Vibrio cholerae YaeO is a Structural Homologue of RNA Chaperone Hfq that Inhibits Rho-dependent Transcription Termination by Dissociating its Hexameric State.
J Mol Biol. 2019 Dec 6;431(24):4749-4766. doi: 10.1016/j.jmb.2019.09.019.
6.
Kumar S, Kumar A, Patel AK.
TIM barrel fold and glycan moieties in the structure of ICChI, a protein with chitinase and lysozyme activity.
Phytochemistry, 2020, 170, 112221
7.
Sonani RR, Rastogi RP, Patel SN, Chaubey MG, Singh NK, Gupta GD, Kumar V, Madamwar D.
Phylogenetic and crystallographic analysis of Nostoc phycocyanin having blue-shifted spectral properties.
Sci Rep. 2019 Jul 8;9(1):9863. doi: 10.1038/s41598-019-46288-4.
8.
Agrawal R, Singh R, Kumar A, Kumar A, Makde RD.
Crystal structures of pyrrolidone-carboxylate peptidase I from Deinococcus radiodurans reveal the mechanism of L-pyroglutamate recognition.
Acta Crystallogr D Struct Biol. 2019 Mar 1;75(Pt 3):308-316.
Chakravarty D, Bihani SC, Banerjee M, Ballal A.
Novel molecular insights into the anti-oxidative stress response and structure-function of a salt-inducible cyanobacterial Mn-catalase.
Plant Cell Environ. 2019 Apr 16. doi: 10.1111/pce.13563.
11.
Gatreddi S, Pillalamarri V, Vasudevan D, Addlagatta A, Qureshi IA.
Unravelling structural insights of ribokinase from Leishmania donovani.
Int J Biol Macromol. 2019 Jun 3. pii: S0141-8130(19)31946-4. doi: 10.1016/j.ijbiomac.2019.06.001.
12.
Gatreddi S, Are S, Qureshi IA.
Ribokinase from Leishmania donovani: purification, characterization and X-ray crystallographic analysis.
Acta Crystallogr F Struct Biol Commun. 2018 Feb 1;74(Pt 2):99-104. doi:10.1107/S2053230X18000109.
13.
Khanppnavar B, Chatterjee R, Choudhury GB, Datta S.
Genome-wide survey and crystallographic analysis suggests a role for both horizontal gene transfer and duplication in pantothenate biosynthesis pathways.
Biochim Biophys Acta Gen Subj. 2019 May 25. pii: S0304-4165(19)30138-2. doi: 10.1016/j.bbagen.2019.05.017.
14.
A conformational switch from a closed apo- to an open holo-form equips the acyl carrier protein for acyl chain accommodation.
Arya R, Sharma B, Dhembla C, Pal RK, Patel AK, Sundd M, Ghosh B, Makde RD, Kundu S.
Biochim Biophys Acta-Proteins Proteom, 186, 163-174 (2019); doi: 10.1016/j.bbapap.2018.12.001.
15.
Carboxypeptidase in prolyl oligopeptidase family: Unique enzyme activation and substrate-screening mechanisms.
Yadav P, Goyal VD, Gaur NK, Kumar A, Gokhale SM, Jamdar SN, Makde RD.
J Biol Chem. 2019 Jan 4;294(1):89-100. doi: 10.1074/jbc.RA118.004254.
16.
Mechanistic basis of vitamin B12 and cobinamide salvaging by the Vibrio species. Agarwal S, Dey S, Ghosh B, Biswas M, Dasgupta J. Biochim Biophys Acta-Proteins Proteom. 2019 Feb;1867(2):140-151. doi: 10.1016/j.bbapap.2018.11.004.
17.
Vibrio cholerae LMWPTP-2 display unique surface charge and grooves around the active site: Indicative of distinctive substrate specificity and scope to design specific inhibitor.
Chatterjee S, Nath S, Ghosh B, Sen U.
Biochim Biophys Acta-Proteins Proteom. 2019 Feb;1867(2):114-124. doi: 10.1016/j.bbapap.2018.11.003.
18.
Gatreddi S, Are S, Qureshi IA.
Ribokinase from Leishmania donovani: purification, characterization and X-ray crystallographic analysis.
Acta Crystallogr F Struct Biol Commun. 2018 Feb 1;74(Pt 2):99-104. doi:10.1107/S2053230X18000109.
1.
Structures and activities of widely conserved small prokaryotic aminopeptidases-P clarify classification of M24B peptidases.
Are VN, Kumar A, Goyal VD, Gotad SS, Ghosh B, Gadre R, Jamdar SN, Makde RD.
Proteins. 2018 Dec 11. doi: 10.1002/prot.25641.
2.
Aggregation-prone Regions in HYPK Help It to Form Sequestration Complex for Toxic Protein Aggregates.
Ghosh DK, Roy A, Ranjan A.
J Mol Biol. 2018 Mar 30;430(7):963-986. doi: 10.1016/j.jmb.2018.02.007.
3.
Crystal structures and biochemical analyses of intermediate cleavage peptidase: role of dynamics in enzymatic function.
Singh R, Goyal VD, Kumar A, Sabharwal NS, Makde RD.
FEBS Lett. 2018 Dec 24. doi: 10.1002/1873-3468.13321.
4.
Structure of Asp-bound peptidase E from Salmonella enterica: Active site at dimer interface illuminates Asp recognition.
Yadav P, Goyal VD, Gaur NK, Kumar A, Gokhale SM, Makde RD.
FEBS Lett. 2018 Oct;592(19):3346-3354. doi: 10.1002/1873-3468.13247.
5.
Structural basis of inactivation of human counterpart of mouse motor neuron degeneration 2 mutant in serine protease HtrA2.
Wagh AR, Bose K.
Biosci Rep. 2018 Oct 5;38(5). doi: 10.1042/BSR20181072.
6.
Tripeptides derived from reactive centre loop of potato type II protease inhibitors preferentially inhibit midgut proteases of Helicoverpa armigera.
Saikhedkar NS, Joshi RS, Bhoite AS, Mohandasan R, Yadav AK, Fernandes M, Kulkarni KA, Giri AP.
Insect Biochem Mol Biol. 2018 Apr;95:17-25. doi: 10.1016/j.ibmb.2018.02.001
7.
Structural insights into the unique inhibitory mechanism of Kunitz type trypsin inhibitor from Cicer arietinum
Bendre AD, Suresh CG, Shanmugam D, Ramasamy S. L.
J Biomol Struct Dyn. 2018 Nov 24:1-9. doi: 10.1080/07391102.2018.1494633.
8.
Structural insights into the nanomolar affinity of RING E3 ligase ZNRF1 for Ube2N and its functional implications.
Behera AP, Naskar P, Agarwal S, Banka PA, Poddar A, Datta AB.
Biochem J. 2018 May 9;475(9):1569-1582. doi: 10.1042/BCJ20170909.
9.
Crystal structure and substrate specificity of ExoY, a unique T3SS mediated secreted nucleotidyl cyclase toxin from Pseudomonas aeruginosa.
Khanppnavar B, Datta S.
Biochim Biophys Acta Gen Subj. 2018 Sep;1862(9):2090-2103. doi:10.1016/j.bbagen.2018.05.021.
10.
Crystal structure of heat shock protein 15 (Hsp15) from Vibrio cholerae: Novel mode of trimerization and nucleic acid binding properties.
Chowdhury SR, Sen U.
Biochem Biophys Res Commun. 2018 Mar 18;497(4):1076-1081. doi: 10.1016/j.bbrc.2018.02.182.
11.
Dynamics of loops at the substrate entry channel determine the specificity of iridoid synthases.
Sandholu AS, Mohole M, Duax WL, Thulasiram HV, Sengupta D, Kulkarni K.
FEBS Lett. 2018 Aug;592(15):2624-2635. doi: 10.1002/1873-3468.13174.
12.
Structure and function of a highly active Bile Salt Hydrolase (BSH) from Enterococcus faecalis and post-translational processing of BSH enzymes.
Chand D, Panigrahi P, Varshney N, Ramasamy S, Suresh CG.
Biochim Biophys Acta-Proteins Proteom. 2018 Apr;1866(4):507-518. doi: 0.1016/j.bbapap.2018.01.003.
13.
drFrnE represents a hitherto unknown class of eubacterial cytoplasmic disulfide oxido-reductases.
Bihani SC, Panicker L, Rajpurohit YS, Misra HS, Kumar V.
Antioxid Redox Signal. 2018 Feb 1;28(4):296-310. doi: 10.1089/ars.2016.6960.
1.
Crystallization and X-ray diffraction analysis of SpaE, a basal pilus protein from the gut-adapted Lactobacillus rhamnosus GG.
Mishra AK, Megta AK, Palva A, von Ossowski I, Krishnan V.
Acta Crystallogr F Struct Biol Commun. 2017 Jun 1;73(Pt 6):321-327. doi: 10.1107/S2053230X17006963.
2.
Crystal structure of Mycobacterium tuberculosis VapC20 toxin and its interactions with cognate antitoxin, VapB20, suggest a model for toxin-antitoxin assembly.
Deep A, Kaundal S, Agarwal S, Singh R, Thakur KG.
FEBS J. 2017 Dec;284(23):4066-4082. doi: 10.1111/febs.14289.
3.
Crystal structure of a novel prolidase from Deinococcus radiodurans identifies new subfamily of bacterial prolidases.
Are VN, Jamdar SN, Ghosh B, Goyal VD, Kumar A, Neema S, Gadre R, Makde RD.
Proteins. 2017 Dec;85(12):2239-2251. doi:10.1002/prot.25389.
4.
Crystal structure determination and analysis of 11S coconut allergen: Cocosin.
Vajravijayan S, Nandhagopal N, Gunasekaran K.
Mol Immunol. 2017 Dec;92:132-135. doi: 0.1016/j.molimm.2017.10.018.
5.
Structure of the human aminopeptidase XPNPEP3 and comparison of its in vitro activity with Icp55 orthologs: Insights into diverse cellular processes.
Singh R, Jamdar SN, Goyal VD, Kumar A, Ghosh B, Makde RD.
6.
J Biol Chem. 2017 Jun 16;292(24):10035-10047. doi: 10.1074/jbc.M117.783357.
Crystal structures reveal N-terminal Domain of Arabidopsis thaliana ClpD to be highly divergent from that of ClpC1.
7.
Mohapatra C, Kumar Jagdev M, Vasudevan D.
Sci Rep. 2017 Mar 13;7:44366. doi: 10.1038/srep44366.
Agarwal S, Dey S, Ghosh B, Biswas M, Dasgupta J. Structure and dynamics of Type III periplasmic proteins VcFhuD and VcHutB reveal molecular basis of their distinctive ligand binding properties.
Sci Rep. 2017 Feb 20;7:42812. doi: 10.1038/srep42812.
8.
Crystal structure and biochemical investigations reveal novel mode of substrate selectivity and illuminate substrate inhibition and allostericity in a subfamily of Xaa-Pro dipeptidases.
Are VN, Kumar A, Kumar S, Goyal VD, Ghosh B, Bhatnagar D, Jamdar SN, Makde RD.
Biochim Biophys Acta. 2017 Feb;1865(2):153-164. doi:10.1016/j.bbapap.2016.10.016.
9.
Mixed-Stack Charge Transfer Crystals of Pillar[5]quinone and Tetrathiafulvalene Exhibiting Ferroelectric Features.
Shivakumar KI, Swathi K, Patil G, Das T, Kumar A, Makde RD, Vanka K, Babu SS, Narayan KS, Sanjayan GJ.
Chemistry. 2017 Jun 29. doi: 10.1002/chem.201702577.
1.
Structural analysis of a penicillin V acylase from Pectobacterium atrosepticum confirms the importance of two Trp residues for activity and specificity.
Avinash VS, Panigrahi P, Chand D, Pundle A, Suresh CG, Ramasamy S.
J Struct Biol. 2016 Feb;193(2):85-94. doi: 10.1016/j.jsb.2015.12.008.
2.
Gating role of His 72 in TmPurL enzyme uncovered by structural analyses and molecular dynamics simulations.
Ghosh B, Goyal VD.
Bioorg Med Chem Lett. 2016 Dec 1;26(23):5644-5649. doi: 10.1016/j.bmcl.2016.10.070.
3.
Crystal structure of the N-terminal domain of human SIRT7 reveals a three-helical domain architecture.
Priyanka A, Solanki V, Parkesh R, Thakur KG.
Proteins. 2016 Oct;84(10):1558-63. doi: 10.1002/prot.25085.
4.
Crystal structure analysis of C-phycoerythrin from marine cyanobacterium Phormidium sp.
A09DM.Kumar V, Sonani RR, Sharma M, Gupta GD, Madamwar D.
Photosynth Res. 2016 Jul;129(1):17-28. doi: 10.1007/s11120-016-0259-5.
5.
Transition of phosphopantetheine adenylyltransferase from catalytic to allosteric state is characterized by ternary complex formation in Pseudomonas aeruginosa.
Chatterjee R, Mondal A, Basu A, Datta S.
Biochim Biophys Acta. 2016 Jul;1864(7):773-86. doi: 10.1016/j.bbapap.2016.03.018.
6.
Crystal structure and dynamics of Spt16N-domain of FACT complex from Cicer arietinum.
Are VN, Ghosh B, Kumar A, Gadre R, Makde RD.
Int J Biol Macromol. 2016 Jul;88:36-43. doi: 10.1016/j.ijbiomac.2016.03.029.
7.
Protein crystallography beamline (PX-BL21) at Indus-2 synchrotron.
Kumar A, Ghosh B, Poswal HK, Pandey KK; Jagannath, Hosur MV, Dwivedi A, Makde RD, Sharma SM.
J Synchrotron Radiat. 2016 Mar;23(2):629-34. doi:10.1107/S160057751600076X.
8.
KatB, a cyanobacterial Mn-catalase with unique active site configuration: Implications for enzyme function.
Bihani SC, Chakravarty D, Ballal A.
Free Radic Biol Med. 2016 Apr;93:118-29. doi: 10.1016/j.freeradbiomed.2016.01.022.
9.
Crystal structure of phosphoglucomutase from Leishmania major at 3.5 Å resolution.
Waugh B, Sen U, Banerjee R. Biochimie. 2016 Feb;121:102-11. doi: 10.1016/j.biochi.2015.11.015.
Science Highlights
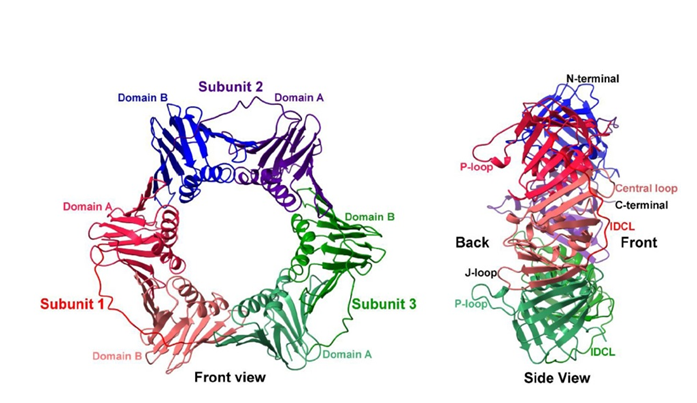
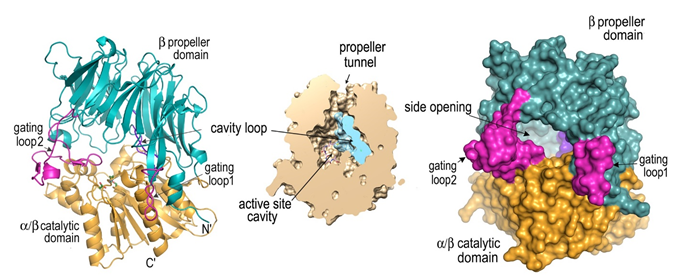
X-ray crystal structure (1.95 Å resolution; PDB id: 7EP8) of Neurospora crassa PCNA (NcPCNA) and compared its structure–function relationship with other available PCNA studies to understand this cross-species incompatibility. Eukaryotic proliferating cell nuclear antigen (PCNA) plays an essential role in orchestrating assembly of the replisome complex, stimulating processive DNA synthesis, and recruiting other regulatory proteins during the DNA damage response. (Ref: Kumari et al, J.Biol Chem, 2021; DOI: 10.1016/j.jbc.2021.100911)
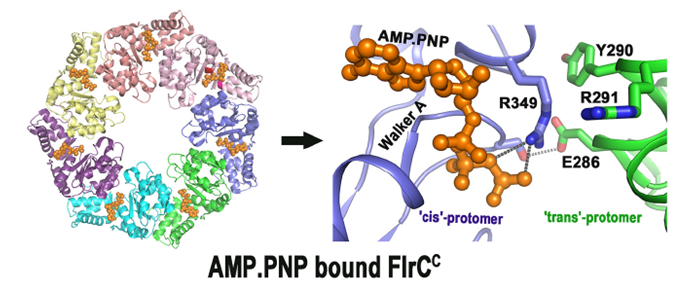
FlrC protein controls motility and colonization of Cholera bacteria for its pathogenesis. Crystal structures of the FlrC mutant proteins determined at BL21 shed light on the mechanism of the colonization. (PDB id: 6LUF, 6LUA) Ref: Chakraborty et al, J Biol Chem, 2020
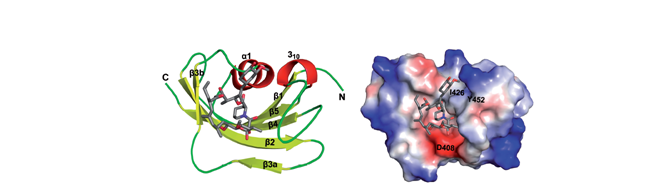
Crystal structure of AtFKBP53 C-terminal domain (Arabidopsis thaliana) at 1.13 Å. Sequence alignment and crystal structure of AtFKBP53 CTD reveals a canonical FKBD fold. The C-terminal domain showed strong PPIase activity, no role in histone chaperoning and revealed a monomeric five-beta palm-like fold that wrapped over a helix, typical of an FK506-binding domain. (PDB id: 6J2M) (Singh et al, 2020, Nucleic Acids Res 48: 1531-1550)
Phylogenetic and crystallographic analysis of Nostoc phycocyanin (2.35 Å resolution) having blue-shifted spectral properties. Sonani, et al, (2019) Sci Rep 9: 9863-9863 PDB id: 6JPR
Crystal structures of S9C peptidase from Deinococcus radiodurans exhibits novel substrate screening mechanism in first carboxypeptidase of prolyl oligopeptidase family. (PDB ID: 5YZN, 5YZM, 6IKG, 6IGR, 6IGQ, 6IGP, 5YZO), Ref: Yadav et al, (2019) J Biol Chem 294: 89-100